脉冲时间依赖可塑性
脉冲时间依赖可塑性(Spike-Timing-Dependent Plasticity,简称STDP)是神经科学中一种极其重要的突触可塑性规则。它把突触强度的改变与突触前、后神经元放电的精确时间差联系起来,被认为是大脑进行时序编码、学习和记忆的核心细胞机制。

一、核心思想:时间差决定方向
传统的赫布学习法则(Hebb’s rule)可以概括为“一起放电的神经元会连接在一起”,但它没有说明放电顺序的重要性。STDP 则进一步细化了这个规则:
- 突触前神经元先放电,突触后神经元紧接着放电(pre-post 模式):突触连接增强,产生长时程增强(LTP)。
- 突触后神经元先放电,紧接着突触前神经元才放电(post-pre 模式):突触连接减弱,产生长时程抑制(LTD)。
也就是说,突触权重如何变化,不只看两个神经元是否都放电了,更取决于谁先放电以及时间差的大小。
下图可以帮助你直观理解这条不对称曲线(用文字表示):
权重变化 Δw
↑
| ● (LTP)
| / \
| / \
|-----/-----\------------→ 时间差 Δt = t_post - t_pre
| / \
| / ● (LTD)
|
- 横轴为时间差 ( \Delta t = t_{post} - t_{pre} )。
- 当 ( \Delta t > 0 )(即突触前先于突触后),权重增加。
- 当 ( \Delta t < 0 )(突触后先于突触前),权重减少。
- 时间差越接近零,变化幅度越大;随着时间差增大,效应呈指数衰减。典型时间窗口约在 ±20~50 毫秒内。
二、发现历史
STDP 现象在 1994-1998 年间被系统性地确立。
- 1994 年,Henry Markram 等人在新皮层锥体神经元之间的突触上,发现重复配对突触前与突触后放电,当突触前放电比突触后放电早约 10 毫秒时,突触增强;颠倒顺序则减弱。
- 1998 年,Guo-qiang Bi 和 Mu-ming Poo 在培养的海马神经元中,精确绘制出了上面那种非对称的时间窗口曲线,并将其命名为“Spike-Timing-Dependent Plasticity”,由此 STDP 成为该领域的通用术语。
三、细胞与分子机制:钙信号的“金凤花原理”
STDP 的极性(LTP 还是 LTD)主要由突触后细胞内钙离子(Ca²⁺)浓度上升的幅度和动力学决定。这里的关键是“不太低也不太高”的浓度诱发 LTD,而快速、大幅的钙浓度升高则诱发 LTP。
1. 基本分子通路
- NMDA 受体:作为重要的钙通道,它的激活需要两个条件同时满足——突触前释放的谷氨酸与突触后膜的去极化(以解除镁离子阻断)。
- 反向传播动作电位(bAP):当突触后神经元产生动作电位时,它不仅沿轴突正向传导,还会反向传播到树突,给树突局部带来大幅去极化。
2. Pre-Post 导致 LTP(Δt > 0)
- 突触前脉冲释放谷氨酸,结合突触后 AMPA 和 NMDA 受体。
- 紧接着(几毫秒内),突触后神经元发放,反向传播的动作电位到达同一突触所在树突分支,去极化强烈,NMDA 受体大量打开,钙离子大量涌入。
- 高浓度的钙激活如 CaMKII(钙/钙调蛋白依赖性蛋白激酶 II)等激酶,引发 AMPA 受体磷酸化并上膜,突触效能增强。这就是“因果性”联结的强化——因为突触前活动恰好“导致”了突触后放电。
3. Post-Pre 导致 LTD(Δt < 0)
- 突触后动作电位先发生,bAP 去极化后,钙水平略有升高,但此时 NMDA 受体并未被谷氨酸充分激活。
- 当突触前脉冲随后到来时,突触后膜的去极化已大幅衰减,NMDA 受体开放程度有限,钙内流只是中等幅度,甚至部分依赖电压门控钙通道或代谢型谷氨酸受体(mGluR)触发的钙释放。
- 这种中等浓度钙优先激活**钙调神经磷酸酶(calcineurin)**等磷酸酶,使 AMPA 受体去磷酸化并内吞,导致突触减弱。
- 在有些突触中(如皮层-纹状体),post-pre LTD 还需要内源性大麻素(eCB)逆行信号参与:突触后去极化产生 eCB,逆行作用于突触前,抑制递质释放。
简而言之:钙峰的高度和时程决定了突触是走向增强还是减弱。
四、数学模型:定量描述时间窗口
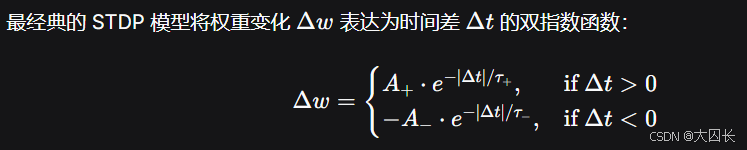
- ( A_+ ) 和 ( A_- ) 分别是 LTP 和 LTD 的最大变化幅度(通常 ( A_- > A_+ ),但 LTD 窗口也更宽,以维持系统稳定)。
- ( \tau_+ ) 和 ( \tau_- ) 是时间常数,通常在 10~30 毫秒。
- 实际应用时,常会加入权重的软、硬边界(如 ( w_{min} \leq w \leq w_{max} )),防止无限增长或降低。
该模型抓住了实验观察到的三点:
- 非对称性:LTP 只对 pre-post 正时差有效,LTD 只对 post-pre 负时差有效。
- 局部性:变化幅度随时间差指数衰减。
- 竞争性:通过 LTD 比 LTP 的窗口稍宽或幅度更大,保证了突触权重的竞争稳定,避免过度兴奋。
五、多样性与调控
STDP 并不是一套死板固定的规则,在不同脑区、不同突触类型、不同调控物质影响下表现出丰富的形式:
- 抑制性突触的 STDP:通常时间窗口对称,或者规律恰好与兴奋性突触相反。
- 对称 STDP:某些突触中,无论 pre-post 还是 post-pre,只要时间差小就增强,时间差大就减弱,形成对称窗口。
- 反赫布 STDP:出现 pre-post 导致 LTD,post-pre 导致 LTP,常见于小脑等结构中。
- 神经调质影响:多巴胺、乙酰胆碱、去甲肾上腺素等可以扩宽或收缩时间窗口,甚至翻转 STDP 极性。这被认为是注意、动机和奖励调控学习的生理基础。
六、功能意义:时序学习与网络自组织
STDP 让神经环路获得了一种对因果性与时序的敏感性,由此支撑了诸多大脑功能:
- 序列学习:在反复经历同一刺激序列时,STDP 会增强从早期事件到后续事件的突触连接,形成一个前馈的“链”,使得网络能学会预测即将发生的事。
- 发育期环路精炼:刚出生时,感觉通路的自发波和感觉传入通过 STDP 精确地修剪突触,形成拓扑投射和感受野。
- 时间差异编码:例如听觉脑干的“计算”双耳时间差来定位声源,其精细延时回路可通过 STDP 进行校准。
- 记忆与海马重放:清醒或睡眠时,海马位置细胞的顺序性放电(sharp wave ripple 中的序列重放)可能通过 STDP 将经验巩固到皮层。
- 预测编码:有“预测”性质的 pre-post 配对被强化,而“意料之外”的 post-pre 配对(先预测到后放电,再收到实际刺激)被削弱,网络可逐步减小预测误差。
七、在计算神经科学和人工智能中的应用
由于 STDP 规则天然无需外部教师信号,只需局部脉冲时序,因此成为**脉冲神经网络(SNN)**最经典的无监督学习算法。
- 特征提取:MNIST 等视觉任务中,两层 SNN 用 STDP 能自主学习出边缘、方向等特征。
- 竞争与平衡:加入侧抑制后,STDP 能驱动不同神经元学习不同模式,实现类似 BCM 理论的发育式学习。
- 神经形态硬件:像英特尔的 Loihi、IBM 的 TrueNorth 等芯片直接在硬件上实现 STDP,用于超低功耗的在线学习。
总结
脉冲时间依赖可塑性把微观脉冲时序精巧地转化为宏观的连接强度变化。它不只是一个细胞生理现象,更是一条普遍的、跨越脑区和物种的学习法则。从海马到皮层、从发育到成年、从感觉编码到高级认知,STDP 都在无声地执行着“因果相连,时序相序”的编码原则,帮助我们的大脑构建起一个既精确又灵活的时序世界模型。

AtomGit 是由开放原子开源基金会联合 CSDN 等生态伙伴共同推出的新一代开源与人工智能协作平台。平台坚持“开放、中立、公益”的理念,把代码托管、模型共享、数据集托管、智能体开发体验和算力服务整合在一起,为开发者提供从开发、训练到部署的一站式体验。
更多推荐

 6
6 0
0- 0



 已为社区贡献26条内容
已为社区贡献26条内容

所有评论(0)