ChIP-seq项目文章 | Nat Commun重庆大学团队发现转录因子Hif1a在皮肤类器官再生毛囊的机制
类器官究竟依靠什么规则,从细胞团块一步步建立出复杂的组织结构?在皮肤类器官领域,已有研究证明分散的表皮细胞和真皮细胞可以在体外重新组装,并形成具备毛囊再生潜力的皮肤样结构。然而,从早期聚合形成的球状体,到后期具有明确表皮—真皮分层的平面化皮肤结构,这一关键形态转变背后的驱动机制并不清楚。
近期,重庆大学雷明星教授团队联合美国南加州大学钟正明院士团队、陆军军医大学烧伤研究所罗高兴教授团队在国际知名期刊Nature Communications发表题为“Metabolic adaptation drives self-organization during skin organoid morphogenesis”的研究论文。该研究以皮肤类器官为研究模型,整合了scRNA-seq、ChIP-seq、ATAC-seq、RNA-seq等多组学和分子功能实验,揭示了低氧如何驱动皮肤类器官从球状聚合体向平面化皮肤结构转变。爱基百客为该研究提供ChIP-seq的技术支持。

研究技术
scRNA-seq、RNA-seq、ATAC-seq、ChIP-seq(Hif1a)、代谢组学等;
整篇研究的思路可以说是用多组学“找机制”,用遗传和药理“做因果”,再用移植实验“看功能结果”。
研究结果
1. 平面皮肤形成过程中缺氧环境的生成
研究发现在air-liquid interface培养中,表皮和真皮细胞会从早期(D4,第4天)球状聚集体,逐渐重组为具有清晰分层的平面化皮肤结构(D7,第7天)。到了D7,类器官不仅形成了类似真实皮肤的表皮-真皮分层和基底膜,还表现出更强的毛囊再生能力。
利用RNA-seq分析,研究发现平面化阶段皮肤类器官中HIF-1信号通路显著激活,而单细胞转录组结果进一步提示表皮细胞富集糖酵解相关基因,表明低氧适应性代谢可能参与了该过程。低氧探针检测证实,类器官培养过程中存在明显的氧梯度:球状阶段表现为球体内部低氧,平面化阶段表现为液相侧低氧、气相侧富氧;同时,从真皮层到表皮基底层及上基层,低氧程度逐渐增强。上述结果提示,差异性的低氧微环境可能是驱动表皮和真皮细胞空间重组及皮肤类器官平面化形成的重要机制。
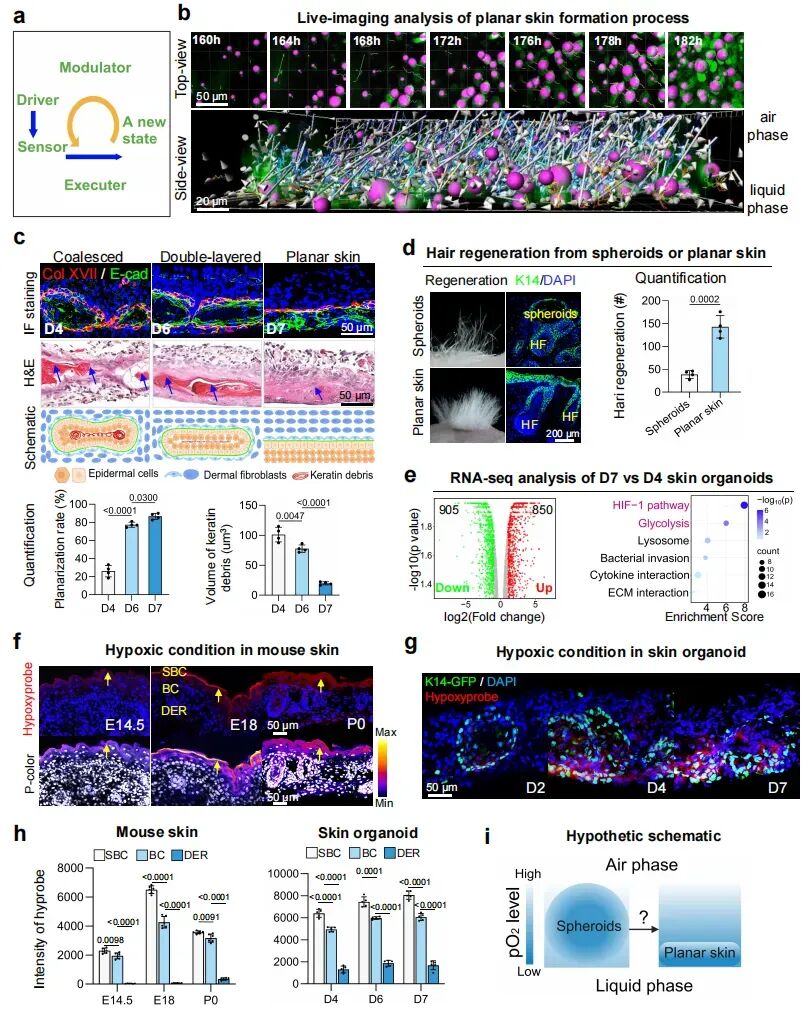
图1:皮肤类器官形态发生过程中的缺氧适应
2. 多组学分析揭示皮肤类器官平面化过程中表皮细胞缺氧信号传导增强
RT-qPCR和ATAC-seq证实,类器官在向平面化结构演变(D7)时,经历了显著的表观遗传重塑与转录重编程,其中Hif1a染色质可及性增加,且其转录活性在众多调控因子中占据主导地位。为了解皮肤球状体和平面皮肤在缺氧条件下的细胞异质性,研究还对D4和D7的类器官进行scRNA-seq分析。研究鉴定出十六个不同的细胞簇,包括五个成纤维细胞簇和两个表皮细胞簇。研究基于单细胞数据构建了一个“缺氧得分”系统,结果发现表皮细胞缺氧得分高于真皮细胞,且在D7时期,表皮细胞的缺氧得分高于D4时期。进一步将表皮细胞细分,研究发现随着表皮细胞从基底细胞(BC)向更成熟的上基底细胞(SBC1-3)分化,其缺氧得分与Hif1a表达量呈同步递增趋势。这部分的研究结果显示Hif1a参与了推动类器官从聚集向平面化的转化过程。
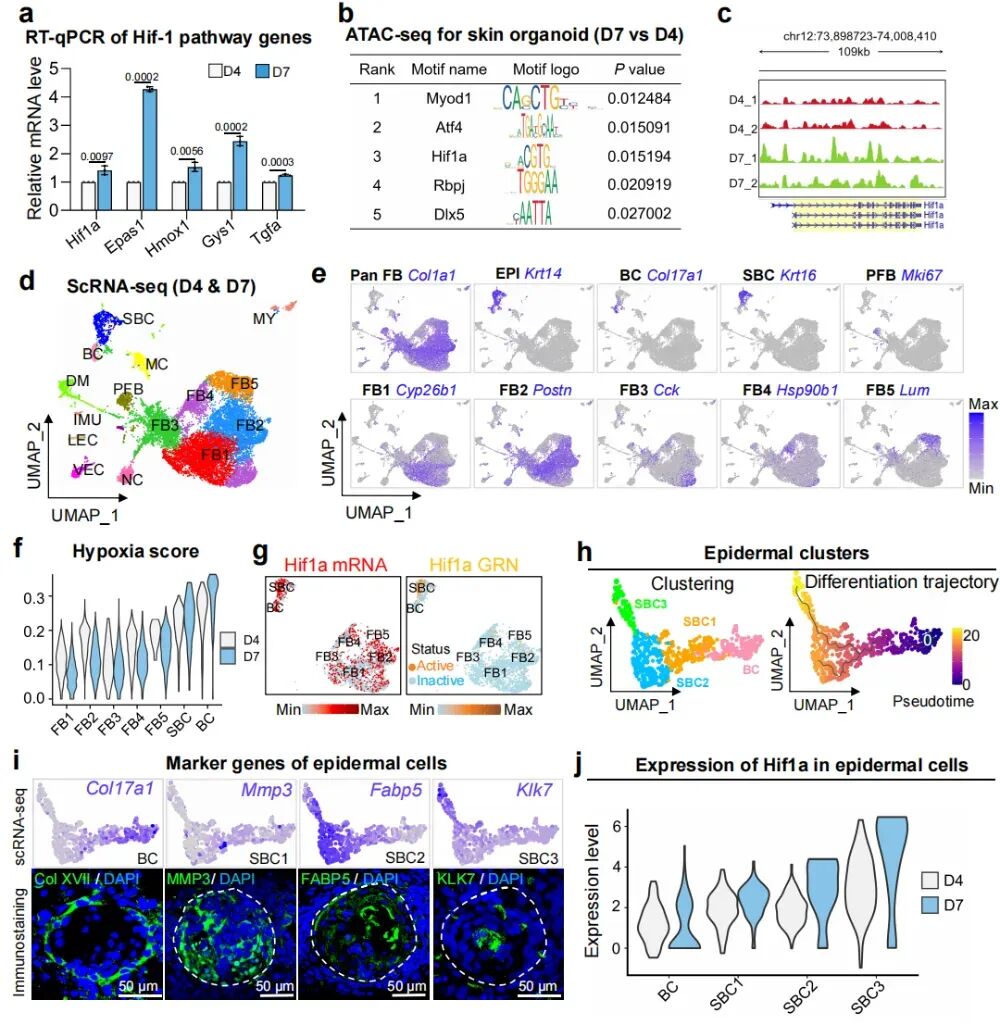
图2:多组学分析揭示了(皮肤类器官)从聚集态向平面态转变过程中的细胞与分子变化。
3. Hif1a驱动皮肤类器官平面化过程中表皮细胞集体的拓扑结构转变
借助RNA-seq、scRNA-seq和免疫染色,研究发现关键并不是HIF2a,而是表皮细胞中的HIF1a。随着培养推进,Hif1a在表皮中持续升高;而抑制HIF1a或在表皮中特异性敲除Hif1a,都会导致类器官平面化受阻,移植后的毛囊再生能力也明显下降。进一步比较不同氧浓度后发现,2%O₂是最适合皮肤类器官自组织的低氧条件,既能促进平面化,也能增强毛囊再生。更关键的是,研究证明真正决定毛囊再生效果的,不是单纯激活HIF1a,而是HIF1a所推动形成了平面化的组织结构本身。
机制上,低氧通过激活HIF1a,增强表皮细胞的糖酵解代谢。针对D4类器官的Hif1a ChIP-seq分析揭示,其结合峰显著富集于HIF-1及糖酵解通路。值得注意的是,多个关键糖酵解酶(如Eno1和Pgam1)被鉴定为HIF-1a的直接靶基因。Motif分析证实,这些结合位点高度富集经典的缺氧响应元件(HRE,RCGTG),且与ATAC-seq鉴定的染色质开放区域高度重合。进一步的启动子序列分析确认,Hif1a是结合于Eno1和Pgam1启动子区排序第一的转录因子。综上所述,HIF-1a通过调节糖酵解促进平面皮肤类器官的自组织,从而增强其毛囊再生的能力。
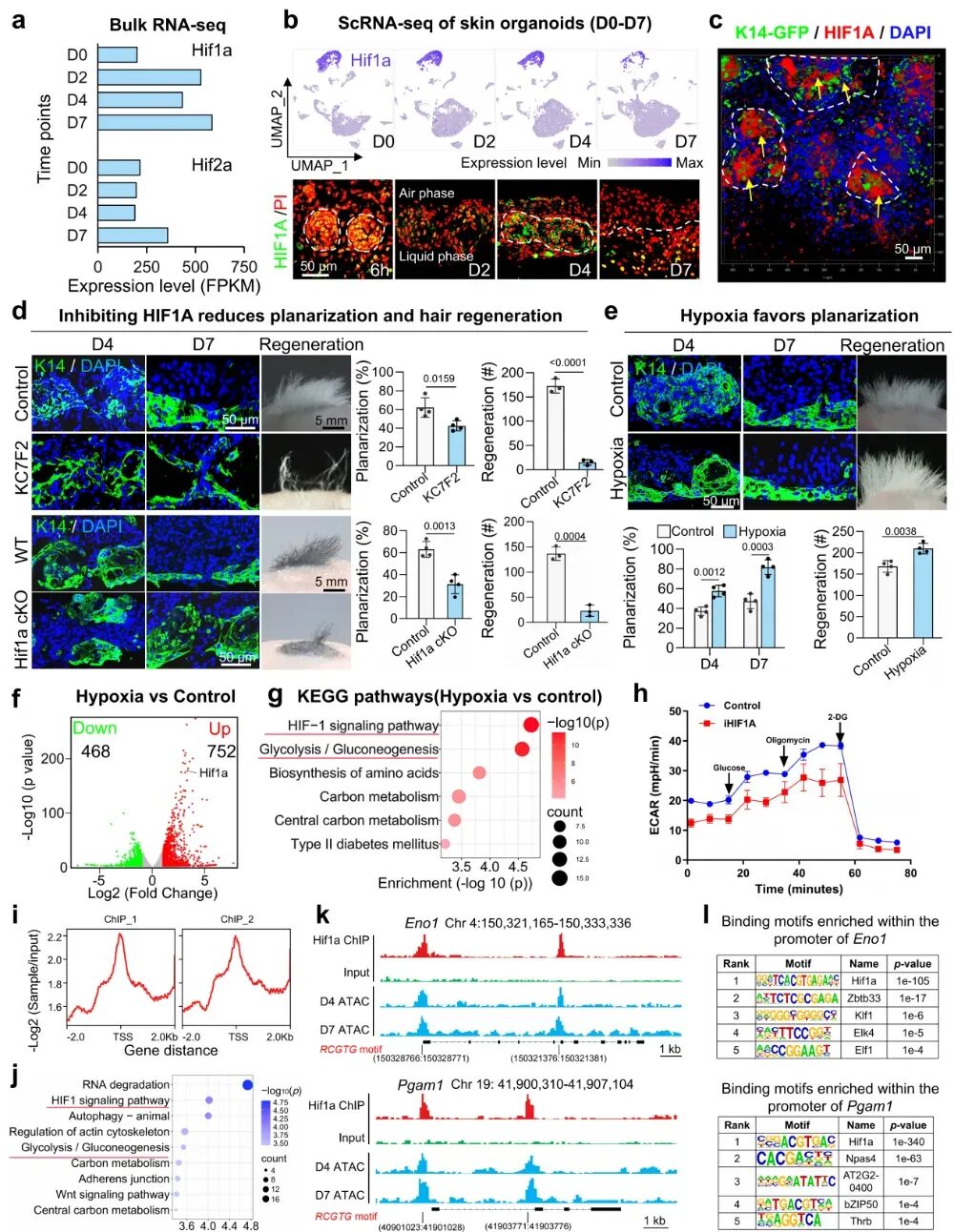
图3:缺氧与Hif1a驱动平面皮肤形成。
4. 需要通过糖酵解过程来驱动平面化过程
研究对D4和D7的类器官进行了代谢组分析,并结合RNA-seq发现D7阶段上调最明显的通路是糖酵解和糖异生。多种糖酵解相关代谢物显著增加,例如G6P、3PG、PEP、乳酸、葡萄糖等。这说明随着类器官从球状结构向平面结构重塑,低氧代谢逐渐增强。为在单细胞水平上解析这一代谢特征,研究基于GSEA数据库构建了糖酵解基因集评分模型。单细胞分析显示SBC和BC的糖酵解评分最高,D7时糖酵解相关基因表达高于D4,乳酸水平在D7也更高。这些说明在平面化过程中,表皮细胞是糖酵解代谢最活跃的细胞群。
研究进一步比较了真皮和表皮的代谢特征,真皮成纤维细胞的氧耗率(OCR)更高,说明它们更依赖氧化磷酸化;表皮角质形成细胞氧耗较低,更偏向糖酵解。线粒体膜电位探针显示真皮细胞氧化代谢更强,pH探针显示表皮细胞酸性信号更高,符合糖酵解活跃的特点。这提示类器官内部形成了明确的代谢分工。真皮细胞主要耗氧,制造低氧微环境;表皮细胞适应低氧,转向糖酵解。研究测了D4和D7的pH,发现范围大约在7.1–7.4,并没有达到典型酸中毒水平(pH<7.0)。所以,这里的表型变化不是由细胞外酸化引起的,而主要是由低氧环境本身驱动的。功能实验用2DG抑制糖酵解后,类器官无法形成平面化皮肤。Hif1a+表皮细胞特异性高表达多种糖酵解关键酶基因:Pkm、Ldha、Pgk1、Eno1、Pgam1。进一步抑制ENO1或PGAM1后,平面化过程被阻断,乳酸和ATP生成下降,移植后的毛囊再生能力下降。这说明糖酵解不仅相关,而且是平面化和后续功能成熟所必需的。
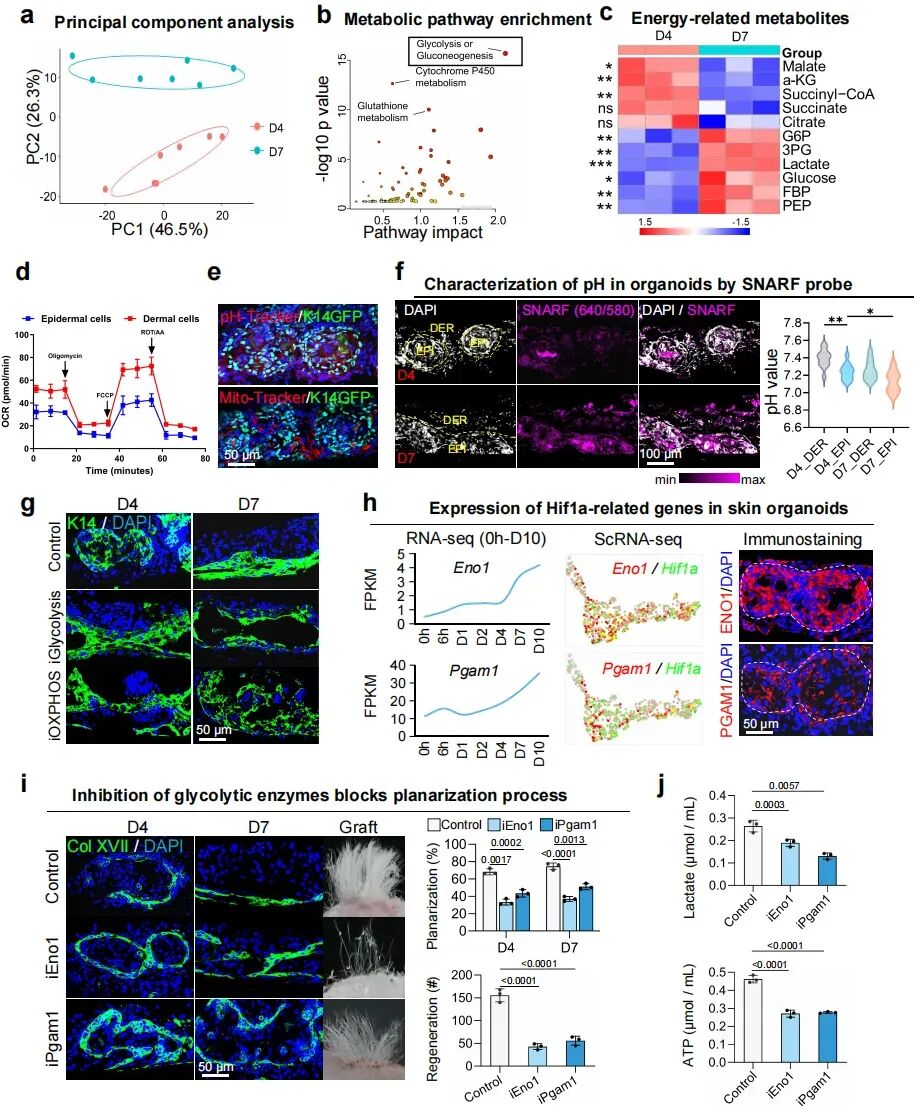
图4:糖酵解代谢促进表皮-真皮相分离。
5. 缺氧可激活溶酶体以水解角蛋白碎片
角质形成细胞先表达高增殖相关角蛋白,随后在上基底层发生凋亡,最终形成位于球体内部的角蛋白碎屑。其中,单细胞分析显示SBC3亚群高表达KLK7(代表表皮分化最末端的细胞状态)。这个细胞群富集于皮肤发育、表皮发育、角质细胞分化,说明这些内层上皮细胞正是角蛋白碎屑的主要来源。RNA-seq和单细胞分析显示,在类器官平面化阶段,溶酶体相关基因显著上调,且表皮细胞具有较高的溶酶体活化评分。这提示表皮细胞除了参与分化,还可能承担了“吞噬清除”的功能。更关键的是,这一清除过程受到低氧直接驱动。
实验表明,低氧培养可显著减少角蛋白碎屑,而抑制HIF1a则会导致碎屑堆积。透射电镜和LysoTracker检测进一步证实,与常氧培养组相比,缺氧培养组的溶酶体信号显著增多。值得注意的是,即使在HEPES缓冲维持pH7.4的条件下,低氧依然能够诱导LAMP2、Ctsl、Ctsd上调,说明这一反应并非继发于酸化,而是由低氧张力本身直接触发。在机制执行层面,研究锁定了溶酶体中的关键水解酶CTSL和CTSD。这两类组织蛋白酶在上基底层表皮细胞中高表达;而当其活性被抑制后,角蛋白碎屑在球体内部显著增加,类器官平面化受阻,移植后的毛囊再生能力也明显下降。这些研究结果表明,缺氧环境可激活溶酶体,从而清除角蛋白碎片。
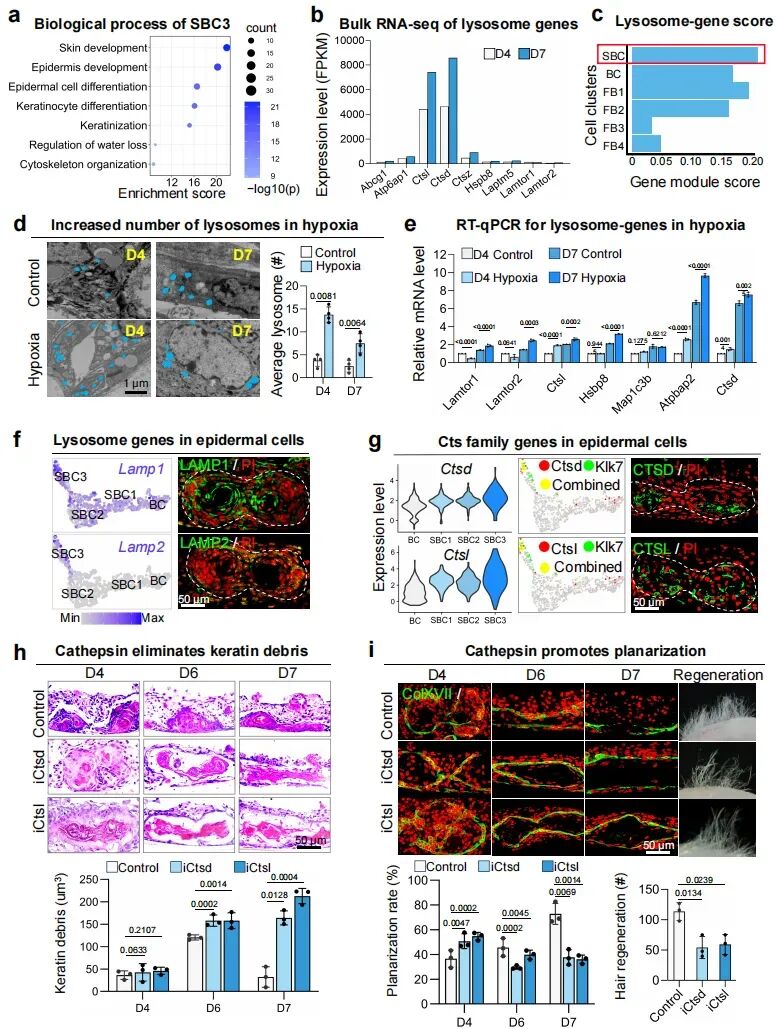
图5:缺氧激活的溶酶体水解酶可清除角蛋白碎片。
6. 缺氧可触发成纤维细胞中异质性的视黄醇代谢
研究发现皮肤类器官中的真皮成纤维细胞可以分成5个亚群(FB1-FB5),其中FB1和FB2靠近表皮,FB3–FB5更远离表皮。进一步分析表明,FB1最可能是乳头层成纤维细胞的前体细胞,而且在D7阶段占比最高。更关键的是,FB1具有非常突出的视黄醇代谢特征。它不仅特异性表达Rara、Rbp4、Rxra、Cyp26b1等基因,还表现出所有真皮亚群中最高的Hif1a水平,提示靠近表皮的低氧微环境可能驱动了这类细胞的代谢重编程。
研究还通过scRNA-seq发现,在小鼠皮肤发育过程中Rara和Rbp4与乳头层标志基因Trps1、Bmp4共表达,并在P0阶段观察到Rara和Rbp4表达上调,说明它们可能同样参与了小鼠乳头状纤维细胞的发育与成熟过程。最后,研究用CellChat和CellCall分析真皮细胞与表皮基底细胞(BC)之间的通讯,发现FB1和BC的互作数量最多、强度最强。这种互作主要依赖层粘连蛋白通路,这很关键,因为层粘连蛋白是基底膜的重要组成部分。所以研究提出FB1不仅在分子特征上最像乳头层成纤维细胞,还可能通过层粘连蛋白相关的ECM-受体互作,与表皮基底细胞共同建立表皮-真皮连接和基底膜结构,从而决定其空间定位并维持皮肤稳定性。
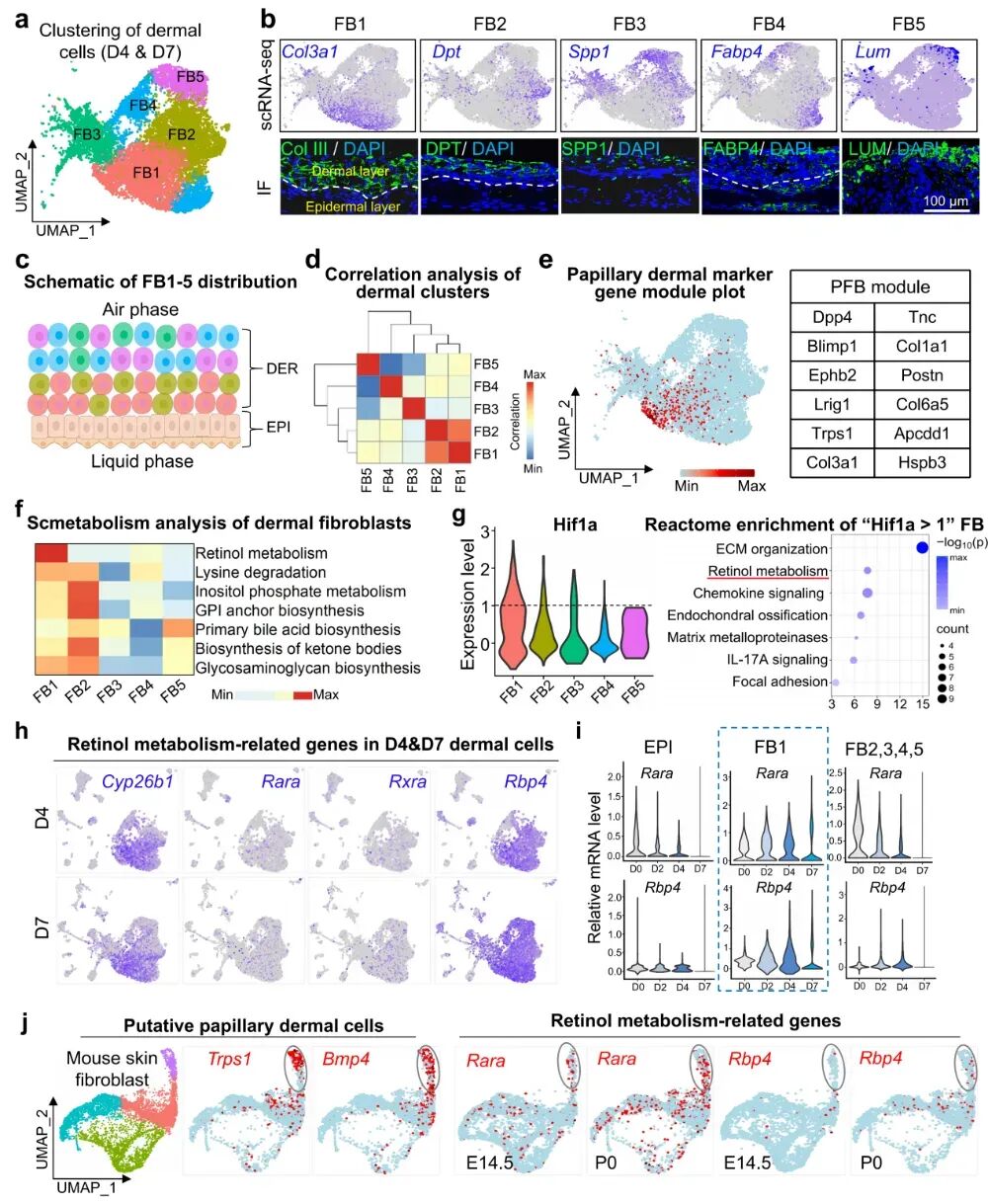
图6:真皮成纤维细胞的代谢异质性。
研究发现视黄醇相关分子RARA和RBP4在表皮-真皮连接处明显富集,尤其是RBP4表达最强。抑制RBP4后,表皮-真皮连接处的胶原合成减少,基底膜稳定性受损;而外源补充视黄酸则可以逆转这一过程。功能上,这条通路同样与皮肤器官成熟密切相关。研究显示,抑制视黄醇代谢会损害皮肤附属器形成,并使移植后毛囊再生能力下降约51%。相反,外源加入全反式视黄酸(atRA)可增强表皮细胞增殖,并减少角蛋白碎屑堆积,提示视黄酸不仅参与组织连接,也有助于表皮更新与结构成熟。
更有意思的是,研究进一步证明,真皮信号还能反向调控表皮代谢。RT-qPCR显示,atRA可上调表皮中的Aldh3b1和Ldha,而抑制RBP4会下调Aldh3b1和Gpi。其中Ldha对低氧条件下维持NAD+再生至关重要,说明真皮来源的维甲酸代谢物并不是简单促进分化,而是可能通过选择性调节糖酵解关键节点,精细控制表皮细胞的能量生成。
因此,这项研究最终提出了一个完整的形态发生模型:缺氧作为驱动力,激活HIF1a作为感受器;表皮细胞通过糖酵解适应低氧,真皮成纤维细胞通过视黄醇代谢重塑微环境,二者共同推动皮肤类器官组装成平面化皮肤结构。
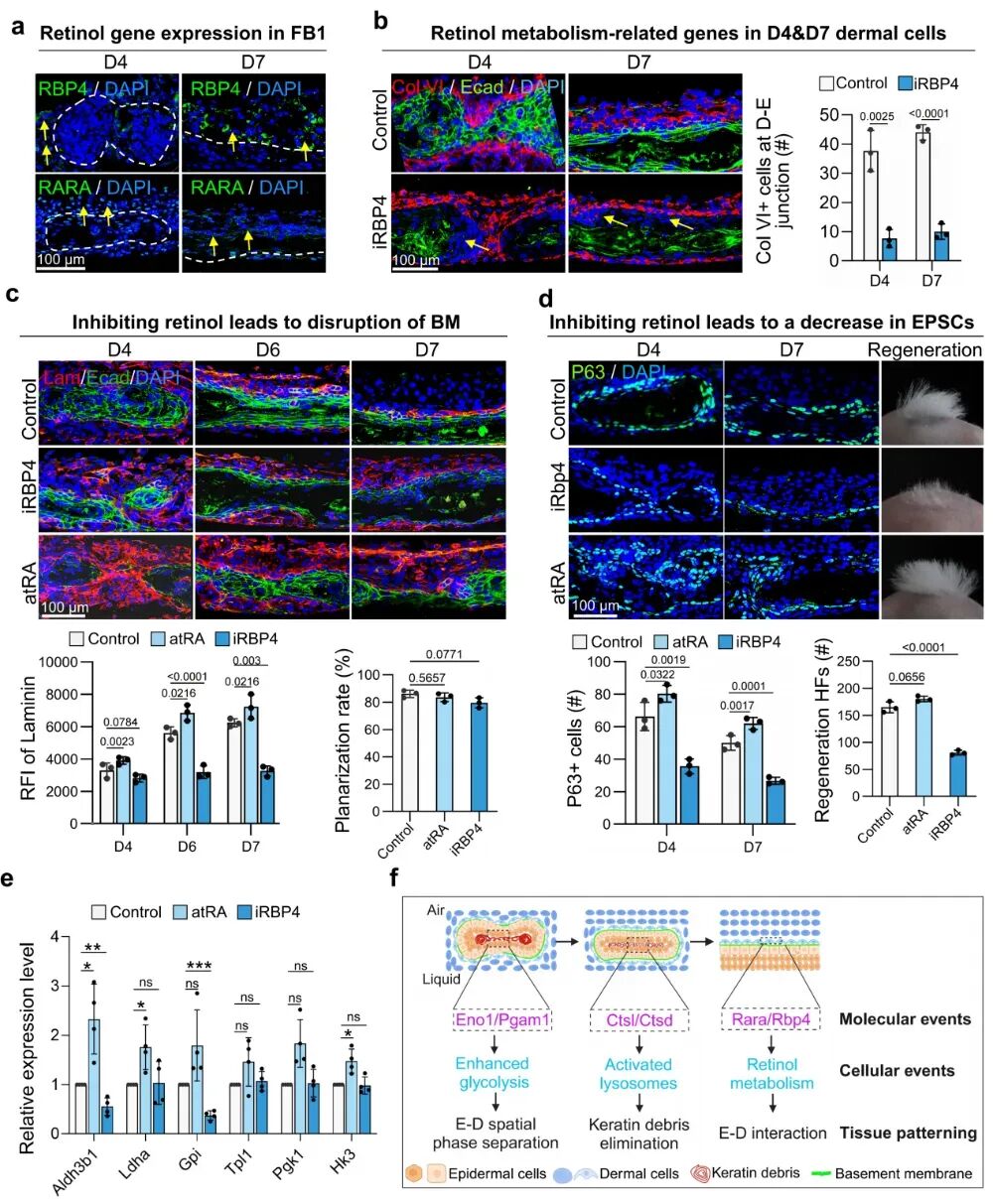
图7:视黄醇代谢支持真皮成纤维细胞的自组织及基底膜形成。
这项研究最重要的价值,在于它把“缺氧”从类器官培养中的一个被动限制因素,重新定义为驱动组织自组织的关键发育信号。研究提出,在皮肤类器官形态发生过程中,低氧并不是简单地造成生长压力,而是通过诱导表皮和真皮细胞产生不同的代谢适应,协调细胞定位、分层、碎屑清除和基底膜重建,最终推动球状结构向平面化皮肤结构转变。
从机制上看,这篇文章揭示了一个清晰的逻辑链:低氧增强→ HIF1a激活→表皮糖酵解上调、溶酶体清除增强,同时真皮成纤维细胞发生视黄醇代谢重编程→最终实现表皮-真皮有序重建。
从意义上看,这项工作不仅加深了我们对皮肤类器官自组织规律的理解,也提示我们在类器官构建中,微环境中的氧和代谢状态,可能和经典形态发生信号一样重要。这对于优化类器官培养体系、提高组织再生能力,以及推动再生医学和皮肤工程应用,都具有重要启发意义。
*本文基于已发表的科学研究进行科普解读,不构成任何医疗建议。*

AtomGit 是由开放原子开源基金会联合 CSDN 等生态伙伴共同推出的新一代开源与人工智能协作平台。平台坚持“开放、中立、公益”的理念,把代码托管、模型共享、数据集托管、智能体开发体验和算力服务整合在一起,为开发者提供从开发、训练到部署的一站式体验。
更多推荐

 11
11 0
0- 0



 已为社区贡献4条内容
已为社区贡献4条内容

所有评论(0)